RNA를 통한 유전자 전사 조절 원리 규명
세포가 어떤 유전자를 얼마나 발현하느냐에 따라 그 세포의 모양, 기능, 수명 등이 결정되므로 유전정보를 처음으로 발현하는 RNA 합성효소의 활성은 세포 내에서 매우 중요하게, 또 정교하게 조절된다. 그러나 이러한 유전자 전사(transcription) 조절의 중요성에도 불구하고 RNA 합성효소가 이러한 단백질과 RNA들에 의해서 어떻게 조절되는지 분자적인 수준에서는 잘 알려져 있지 않았다.
☞ 유전자 전사: DNA의 유전정보가 RNA에 옮겨지는 과정을 말한다. 유전정보의 복사물인 RNA는 단백질 합성에 사용된다.
KAIST는 화학과 강진영 교수 연구팀이 RNA를 통한 RNA 합성효소의 조절 메커니즘을 알아내고자 RNA 합성효소와 RNA 합성효소를 조절하는 바이러스 유래 RNA인 *HK022 putRNA의 결합 구조를 초저온 전자현미경(cryo-EM)으로 규명하여 유전자 전사조절의 기초 원리를 규명했다고 7일 밝혔다.
*HK022 putRNA: HK022 박테리오파지(박테리아를 감염시키는 바이러스)의 RNA로 다른 단백질의 도움 없이 해당 RNA를 만든 RNA 중합효소와 결합해 RNA 합성이 계속 되도록 RNA 중합효소를 조절
KAIST 화학과 황승하 박사과정이 제1 저자로 참여한 이번 연구는 국제 학술지 `네이처 커뮤니케이션즈 (Nature Communications)'에 지난 8월 15일 출판되었다. (논문명: Structural basis of transcriptional regulation by a nascent RNA element, HK022 putRNA, (nature.com)).
HK022 putRNA는 RNA 합성효소와 결합해서 RNA 합성이 멈추지 않고 계속 되도록 도와주는 역할을 한다. 이러한 기능을 이해하기 위해서 본 연구팀은 putRNA와 RNA 합성효소의 결합 복합체(put-associated RNA polymerase elongation complex, putEC)의 세 가지 구조를 초저온 전자현미경으로 규명하였다. (그림 1)
이 연구에서는 활성을 가진 putRNA를 제작하기 위해 장애물 단백질을 RNA 합성에 활용하는 방법을 고안하였으며, 초저온 전자현미경 촬영 결과 예상하지 못했던 세 종류의 복합체 – putRNA가 잘 접혀서 RNA 합성효소와 결합하고 있는 putEC, put RNA가 접히지 않은 put-없는 EC, 잘 접힌 putRNA와 시그마 단백질이 함께 RNA 합성효소와 결합하고 있는 시그마* 결합-putEC – 를 발견할 수 있었다. (그림 1)
*시그마: RNA 합성효소가 유전자 RNA 합성을 처음 시작할 때 필요한 단백질로 RNA 합성이 어느 정도 안정화되면 RNA 합성효소에서 떨어진다.
연구팀은 이들 복합체의 구조를 통해 putRNA가 이전 연구에서 예측된 대로 RNA 합성효소와 안정적으로 결합하고 있지만 예측과 달리 예상보다 더 많은 염기쌍(base pair)을 사용해 RNA 이중나선(double helix) 뿐 아니라 삼중나선(triple helix)을 형성하는 것을 확인하였다. 또한, putRNA가 RNA 합성효소와 결합하면 RNA 합성효소가 RNA 합성을 잠시 멈출 때 가지는 구조의 변화를 방해해서 RNA 합성을 지속하도록 한다는 가설을 제시할 수 있었다.
한편, 시그마 단백질(σ70)은 RNA 합성효소가 전사를 시작할 때 필요한 전사 개시인자로, RNA 합성이 안정되면 RNA 합성효소에서 떨어졌다가 특정 DNA 서열(–10-유사 서열)이 있으면 전사 과정 중이라도 다시 RNA 중합효소와 결합해 RNA 합성을 일시적으로 멈추는 것으로 알려져 있다. 이번 연구에서는 예상치 못하게 관찰된 시그마 결합-putEC 구조를 통해 시그마가 RNA 합성효소와 결합하여 RNA 합성이 잠깐 멈추면 putRNA가 더 잘 접힌다는 것을 알 수 있었다.
이 연구의 교신저자인 강진영 교수는 "RNA 합성효소는 세포 내에 저장된 유전 정보를 처음으로 꺼내어 생명활동에 활용하는, 세포 내에서 제일 중요한 단백질 중 하나이다. 그러나 RNA 합성효소의 큰 크기와 다양한 구조 변화 때문에 이전에 주로 활용하던 X-ray 결정학 방식으로는 그 구조를 관찰하기가 어려웠다. 최근 초저온 전자현미경의 발달로 이제야 조금씩 RNA 합성효소의 작동 원리가 알려지고 있는 상황이다. 이번 연구는 이전에 잘 알려지지 않았던, RNA를 통한 전사 조절의 기초적인 원리를 설명한 것으로, RNA를 통한 RNA 합성효소 조절의 다양한 전략을 밝혀줄 시작점이며, 더 나아가 유전자 발현을 조작할 수 있는 RNA의 개발을 도울 수 있는 정보를 제공할 것이라 기대한다.ˮ고 밝혔다.
한편 이번 연구는 한국연구재단의 이공분야기초연구사업(우수신진연구)과 원천기술개발사업의 지원을 받아 수행됐다.

그림 1. 바이러스 유래 RNA, HK022 putRNA와 대장균 RNA중합효소가 결합한 복합체의 초저온 전자현미경 구조. 왼쪽부터 putEC, put-없는 EC, 시그마-결합 putEC
□ 연구개요
1. 연구배경
전사(transcription)는 DNA에 저장된 유전정보를 RNA로 옮기는 생명활동으로 생명을 유지하고 재생산하는데 핵심적인 생화학 반응이다. 대부분의 바이러스는 숙주(host) 세포 내에서 자신의 유전자를 효과적으로 발현하기 위해 숙주의 전사 과정을 조작하는 다양하고도 영리한 전략을 가지고 있다. 1970년대 홍콩에서 발견된 HK022 박테리오파지는 역사적으로 전사 연구에 많이 활용된 람다파지와 같은 속(family)에 속하며 put (polymerase-utilization)이라고 명명된 RNA를 사용하여 숙주의 RNA 중합효소가 자신의 유전체를 전사할 때 전사 중지나 종결이 일어나지 않게 한다. putRNA는 다른 단백질 인자 없이 RNA 만으로 전사 중인 RNA 중합효소와 결합하여 전사를 조절한다는 점에서 특이하고 흥미로운 전자조절인자이다.
HK022 putRNA가 두 개의 스템-루프 (stem-loop)구조를 가지고 숙주인 대장균(E.coli)의 RNA 중합효소(RNA polymerase)의 아연결합부분(zinc binding domain, ZBD)에 결합할 것이라는 가설이 이전의 돌연변이 실험을 통해 제시된 바 있으나 실제로 putRNA와 RNA중합효소 결합 복합체의 3차원 구조는 알려져 있지 않았다. 이에 본 연구팀에서는 HK022 putRNA와 RNA중합효소 복합체를 제작하고 이를 초저온 전자현미경(cryo-EM)으로 관찰하여 HK022 putRNA가 어떻게 전사를 조절하는 기능을 가지는지 이해하고자 하였다.
2. 연구내용
RNA 합성에 흔히 쓰이는 화학적 합성이나 T7 RNA 중합효소에 의한 합성으로는 활성이 있는 putRNA를 제작할 수 없다. 이는 putRNA가 기능을 가지기 위해서는 숙주의 RNA 중합효소에 의해 합성되면서 효소와의 상호작용을 통해 특이적인 구조로 접혀야 하기 때문이다. 따라서 본 연구실에서는 실제 세포 내의 환경과 유사하게 대장균 RNA 중합효소와 전시개시인자인 시그마 인자(σ70)를 사용하여 putRNA를 합성한 후 합성된 RNA와 이를 합성한 RNA 중합효소의 복합체인 putEC를 급속 냉각하여 이를 초저온 전자현미경으로 관찰하는 방법을 고안하였다. 이 때, 일정한 길이의 RNA를 합성하기 위해 RNA 합성에 활용할 DNA에 Lac 리프레서 (LacI) 단백질이 결합하는 서열을 삽입하고 LacI를 첨가하여 RNA중합효소가 putRNA를 합성한 후 특정 위치에서 LacI에 막혀서 더 이상 RNA 합성을 하지 못하도록 하였다. (그림1)
cryo-EM 데이터 분석을 통해 시료 내에 putRNA가 잘 접혀져서 RNA 중합효소와 결합하고 있는 putEC, putRNA가 접히지 않은 put-less EC, putRNA가 잘 접혀 있으며 시그마인자도 RNA중합효소와 결합하고 있는 시그마-결합 putEC, 총 세 종류의 복합체가 존재하는 것을 확인할 수 있었다. (그림 2)
putEC 구조에서 putRNA는 두 개의 스템-루프(stem-loop) 구조를 V-자 형태로 이루어 기존 연구에서 제시되었던, RNA중합효소의 아연결합부분(β’zinc-binding domain, β’ZBD)과 결합하는 것을 확인하였다. 한편, 기존 연구에서 예측한 것과 다른 점은 (1) putRNA의 접힘 구조에 63 개가 아닌 70 개의 염기들이 참여하고 있고, (2) putRNA의 기능에 중요한 35번 구아닌 염기가 단독으로 존재하지 않고 9번 아데닌과 염기쌍을 이루고 있으며, (3) 첫 번째 스템-루프 구조는 이중나선이 아닌 삼중나선 구조를 형성하고 있었다는 점이다. (그림 3) 이 세 가지 구조적 특징을 제거하는 돌연변이를 putRNA에 삽입하면 putRNA의 기능이 줄어드는 것을 통해 위 세 가지 특징이 putRNA의 구조 형성과 기능에 중요하다는 것을 확인하였다.
또한, putEC와 put-less EC의 구조 비교를 통해서 putRNA가 RNA중합효소가 정지할 때 가지는 구조 변화를 막는 것이 관측되었다. RNA 합성을 멈춘 상태의 RNA중합효소들은 공통적으로 효소의 일부분이 최대 5°까지 돌아가는, ‘회전 (swiveling)’이라고 알려진 구조 변화를 가지는데, putRNA를 가지는 putEC의 RNA중합효소는는 putRNA가 없는 put-less EC의 효소보다 더 작은 ‘회전’ 각도를 보였다. (그림 4) 따라서 putRNA는 전사 정지를 일으키는 ‘회전’을 저해하여 전사 정지를 줄이는 것으로 보인다.
한편, 시그마 인자는 전사가 시작될 때 RNA중합효소와 결합하여 프로모터DNA를 인식하도록 도와주는 전사 개시인자로 중합효소가 프로모터DNA를 지나 안정적인 전사 연장단계에 도달하면 RNA중합효소에서 떨어져 나간다. 그러나, DNA에 프로모터 서열과 유사한 ‘-10-유사서열 (-10-like element)’이 존재하면 이 위치에 시그마인자가 다시 결합하여 RNA중합효소의 전사 정지를 일으킨다는 사실이 알려져 있다. putRNA는 23-28번위치에 ‘-10-유사서열’을 가지고 있는데, 본 연구에서 발견된 ‘시그마-결합 putEC’는 이 ‘-10-유사서열’을 통해 시그마가 RNA중합효소와 재결합한 복합체로 보인다. 흥미롭게도 시그마가 없는 RNA중합효소 중 반 정도만이 잘 접힌 putRNA를 가지고 있는데 반해, 시그마가 결합된 RNA중합효소는 대부분 잘 접혀있는 putRNA를 가지고 있었다. 이는 시그마의 결합과 이로 인한 전사의 일시 중지가 putRNA의 접힘을 돕는 효과를 가지는 것을 의미한다. (그림 5) 여러 종의 putRNA들이 ‘-10-유사서열’을 포함하는 것으로 보아 ‘-10-유사서열’로 인해 일어나는 ‘시그마-의존 전사 정지’가 putRNA의 합성에 중요한 것으로 보인다. 또한, 시그마가 결합된 RNA 중합효소는 LacI 단백질에 의해 전사를 멈추지 않고 더 진행하여 더 긴 길이의 RNA를 합성하는 것이 관측되었는데, 이는 연장 단계에서 시그마 인자가 결합이 우리의 예상보다 더 많은 전사 조절 역할을 가지고 있음을 시사한다.
3. 기대 효과
본 연구에서 연구팀은 전사 조절 RNA인자인 HK022 put이 RNA중합효소와 결합한 전사 복합체의 구조를 초저온 전자현미경으로 규명하여 put의 전사 조절 기작을 밝혔다. 구조를 통해 putRNA가 RNA중합효소의 ‘회전’ 구조 변화를 막아 전사 정지를 방해하는 것을 관찰하였으며, ‘-10-유사 서열’에 의한 시그마-의존 전사 정지가 putRNA의 접힘을 돕는다는 가설을 제시하였다. 본 연구 결과는 생명활동에 매우 필수적이면서 기초적인 전사 과정이 RNA에 의해 조절되는 분자적 원리를 설명하는 연구로 더 나아가 앞으로 다양하게 이루어질 핵산의 전사 조절 역할을 이해하는 데 필요한 구조적 기반을 제시할 것으로 기대한다.

그림 1. putEC의 제작 과정. RNA합 성에 필요한 프로모터와 putRNA 서열, LacI 단백질이 결합하는 lacO 서열을 포함하는 DNA 스캐폴드에 RNA중합효소와 시그마 인자, LacI를 첨가한 후 rNTP를 넣어 putRNA- RNA중합효소 복합체(putEC)를 제작한다. 이후 남은 rNTP를 제거하고 IPTG를 첨가하여 LacI를 DNA에서 제거한 후 이를 초저온 전자현미경 분석에 사용하였다.

그림 2. 시료에서 발견된 세 가지 다른 형태의 RNA중합효소 복합체 구조. 왼쪽부터 putEC, put-less EC, 시그마-결합 putEC이다.

그림 3. putRNA의 구조 비교. (왼쪽) 이전에 계산을 통해 예측된 구조 (King et al., 1996), (오른쪽) 초저온전자현미경으로 규명된 구조.

그림 4. 여러 대장균 RNA중합효소 연장복합체의 ‘회전’ 비교. 비정지 (회색), putEC (파란색), put-less EC (빨간색), 백트랙 정지 (노란색), his 헤어핀 정지 (분홍색) 상태의 RNA중합효소 구조들을 효소 중심 부분을 기준으로 겹친 후 회전하는 모듈의 회전 각도를 측정하였다. (왼쪽) 중첩된 RNA중합효소 구조들의 정면 모습. (오른쪽) 중첩된 RNA중합효소 중 회전 모듈만 표시한 그림. 위에서 본 모습. 측정한 회전 각도가 화살표 옆에 적혀 있다.
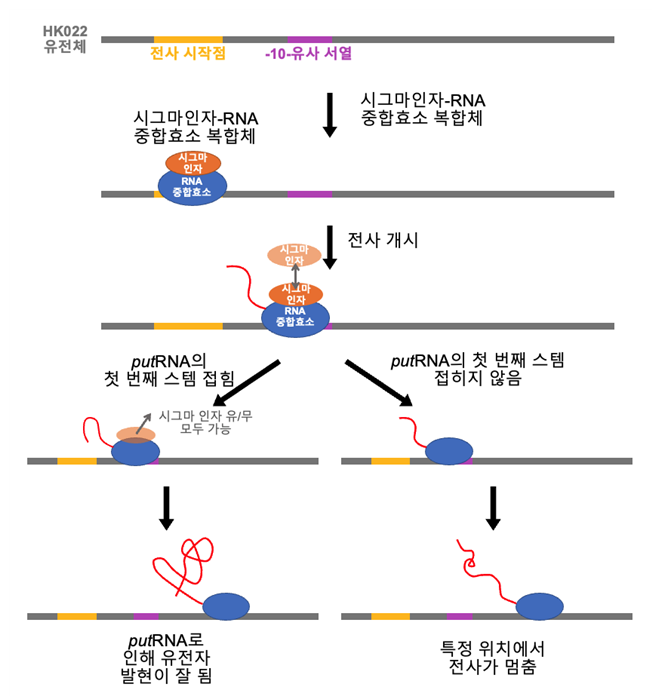
그림 5. HK022 putRNA의 작동 원리. 숙주의 RNA중합효소는 HK022 유전체의 프로모터 부분을 인식하여 putRNA를 합성하기 시작한다. 이때 putRNA가 만들어 지면 RNA중합효소와 결합하여 전사의 중지와 종결을 방해하여 HK022 파지의 유전자를 효율적으로 합성하게 된다. 이 때, putRNA 서열 내부에 있는 –10-유사서열에 시그마 인자가 결합하면 putRNA가 더 잘 접히게 된다. 반면, putRNA가 잘 만들어지지 않으면 이후 전사 종결서열에서 RNA합성이 멈추고 HK022 유전체의 발현이 저해된다.
생명과학 KAIST (2022-09-07)